大脑如何实现灵活应对复杂的世界,整合所有相关信息而不会被无关信息淹没?
神经元的 “混合选择性”
神经元的 “混合选择性” 意味着它们可以参与多个计算而不局限于单一特定功能。这篇论文用混合选择性概念解释了大脑如何选择将相关信息整合在一起而放弃无关信息干扰。
每天,我们周围发生很多事情,我们也有很多内在的动力和记忆,我们的思想必须以某种方式保持既灵活而集中,以指导我们必须做的每件事,我们的大脑都在为此努力优化做出权衡。包括麻省理工学院 Picower 学习与记忆研究所的教授、“混合选择性”(the mixed selectivity )概念的先驱 Earl K. Miller 等在内的一组神经科学家在《神经元》期刊上发表一篇新论文,描述了大脑如何获得认知能力,将所有相关信息整合在一起,而过滤掉无关信息。
作者认为,这种灵活性源于在许多神经元中观察到的一个关键特性:“混合选择性”。虽然许多神经科学家过去认为每个细胞只有一个专用功能,但最近的证据表明,许多神经元可以参与各种计算组合,每个计算组合并行工作。换句话说,当一只兔子考虑在花园里啃一些生菜时,一个神经元可能不仅会评估它有多饿,还会评估它是否能听到头顶上的鹰叫声或闻到树上的土狼的气味,以及生菜有多远。
Earl K. Miller 表示,大脑不能同时处理多项任务,但许多细胞确实有能力被连接到多个计算工作中 (本质上是 “思想”)。在这篇新论文中,作者描述了大脑用来招募神经元进行不同计算的特定机制,并确保这些神经元代表复杂任务的正确维度数量。“这些神经元身兼数职,”“通过混合选择性,你可以得到按实际需求匹配的复杂的代表性空间,而不是更复杂。这就是灵活认知的意义所在。” 研究报告的合著者、索尔克研究所和加州大学圣地亚哥分校的 Kay Tye 教授说,神经元之间的混合选择性,是实现许多心理能力的关键。“混合选择性是赋予我们灵活性、认知能力和创造力的属性。这是最大化计算能力的秘密,而计算能力本质上是智能的基础。”
概念起源
2000 年,Miller 实验室的一项认知研究得出的一个令人惊讶的结果 —— 当动物将图像分类时,大脑前额叶皮层中大约 30% 的神经元似乎参与其中。那些相信每个神经元都有特定功能的怀疑论者嘲笑说,大脑会让这么多细胞只完成一项任务?!Miller 和他的同事 John Duncan 争辩说,也许细胞具有参与许多计算的灵活性,在一个脑力工作小组服务并不妨碍它们为其他许多小组服务。混合选择性的想法由此萌发。
但是混合选择性有什么好处呢?2013 年,Miller 与 IBM 研究院的 Mattia Rigotti 和哥伦比亚大学的 Stefano Fusi(这两位也是新论文的共同作者)合作,展示了混合选择性如何赋予大脑强大的计算灵活性。从本质上讲,具有混合选择性的神经元集合相比具有特定功能的神经元群体,能够容纳更多关于任务的信息维度。Mattia Rigotti 说:“自从我们的原创工作以来,我们通过经典机器学习思想的视角,在理解混合选择性理论方面取得了进展。”“另一方面,实验学家所关心的有关在细胞水平上实现它的机制的问题相比之下还没有得到充分的探索。这次合作和这篇新论文填补了这一空白。
概念具像
并非所有的大脑功能都是复杂的。简单的功能可以通过简单的架构,单层执行。支撑简单功能的架构可具有直接属性、服务于该功能的简单组合,缺乏灵活性能够使得这些功能更快速、高效且模式化。
相比之下,负责复杂思维和行为的神经系统要求灵活性。聪明的思想是灵活的思想。拥有更复杂神经系统的动物可以通过将更多参数整合到决策过程中来改变它们的行为方式。它们会根据当前情况和不断变化的子目标来调整正在进行的行为。它们还会考虑到累积的事件历史,会影响决策阈值。这种概括情境依赖行为的能力,对于预期自己未来的行为、对于制定和执行计划都至关重要。
可以肯定的是,有些神经元专门用于特定的输入 —— 这些细胞具有 “纯选择性”,例如只关注兔子是否看到生菜。一些神经元表现出 “线性混合选择性”,这意味着它们的反应可预测地依赖于多个输入的叠加 (兔子看到生菜就觉得饿了)。增加了最高维度灵活性的神经元是 “非线性混合选择性” 神经元,它们可以解释多个独立变量,可能会权衡一整套独立的条件 (例如,有生菜,我饿了,我没有听到鹰的声音,我没有闻到土狼的气味,但是生菜很远,我看到一个相当坚固的栅栏)。
再例如,想象一只小鼠在其环境中遇到一种没见过的浆果 —— 浆果可能好吃 (这是一个维度),也可能有毒 (那是另一回事),小鼠因而犹豫是否要吃它。然而,如果小鼠随后遇到一个同类,其呼吸带有相同浆果的气味,它可能会将其解释为该浆果可以安全食用的标志 (取决于同伴的表面健康状况)。在这种情况下,小鼠同伴呼吸中的浆果气味可以作为一种社会验证,表明这种食物可能是安全的,值得花功夫去吃。这种社交线索有效地将小鼠对不熟悉浆果的评估从潜在风险转变为可能安全,所有这些都基于食物偏好的社会传播。具有混合选择性的神经系统将能够整合所有这些相关信息。
这种灵活性的神经基质在已知对灵活行为至关重要的皮质区域 —— 如前额皮质 (PFC) 中尤为普遍。前额皮质神经元具有适应性和多元响应特性,即 “混合选择性”。它们身兼数职,在不同的行为背景下表现出不同的选择性模式。信息通常在它们之间广泛传播。前额皮质神经元有中等高的基础放电率,高比例的神经元会对多种刺激做出反应。神经元和它们的轴突提供了各种思想、感觉、感觉和运动指令通行的高速公路。信息载体可以采取不同的路径和不同的目的地。它们可能有时选择这条路径,有时又选择另一条路径。这种自主性提供了最大的自由度和维度。
随之而来的是思想的深度。多元神经元特性可以增加群体的表征维度,允许更复杂的计算。此外,大脑根据手头的任务来调整维度。维度扩展和收缩聚焦处理沿着相关的维度这使处理过程在任务和目标的指导下进行。
简而言之,混合选择性赋予大脑复杂性和灵活性所需的处理能力。这种灵活性的代价是无法利用世界的规律来推广到新的情况。混合选择允许在每个细节中表示大量不同的情况,但有时我们需要丢弃或忽略一些信息以做出正确的决定。
招募神经元
虽然混合选择性有大量的证据支持 —— 在大脑皮层和其他大脑区域,如海马体和杏仁核中都观察到了混合选择性 —— 但仍有悬而未决的问题。例如,神经元是如何被招募到任务中的?如此 “开放” 的神经元是如何只关注对任务真正重要的东西的?在这项新研究中,研究人员定义了观察到的混合选择性的形式,并认为当 “脑电波” 和神经调节剂 (影响神经功能的血清素或多巴胺等化学物质) 将神经元招募到计算集合中时,它们也帮助神经元 “控制” 了什么是重要的。
那么,是什么让神经元集中注意力在显著因素上,不管有多少其他因素?其中一种机制是振荡( oscillations),当许多神经元都以相同的节奏保持它们的电活动时,就会在大脑中产生振荡。这种协调的活动使信息共享成为可能。作者强调的另一种机制是神经调节剂。这些化学物质一旦到达细胞内的受体,也会影响它们的活性。例如,乙酰胆碱的爆发可能同样会使具有正确受体的神经元对某些活动或信息 (比如饥饿感) 进行调谐。
“这两种机制可能一起工作,形成动态的功能网络,” 理解混合选择性对理解认知至关重要。“混合选择性无处不在,从高级认知到物体识别等‘自动’感觉运动过程,它存在于各个物种和各个功能中。混合选择性的广泛存在强调了它在为大脑提供复杂思考和行动所需的可扩展处理能力方面的基本作用。”
参考文献
Mixed selectivity: Cellular computations for complexity
via:
-
大脑如何实现灵活应对复杂的世界,整合所有相关信息而不会被无关信息淹没?生物通 2024 年 05 月 15 日 12:32 广东
注:全文机翻,未校对。
Mixed selectivity: Cellular computations for complexity
混合选择性:复杂性的细胞计算
Kay M. Tye
Earl K. Miller
Felix H. Taschbach
Marcus K. Benna
Mattia Rigotti
Stefano Fusi
Open Access Published:May 09, 2024 DOI:https://doi.org/10.1016/j.neuron.2024.04.017
Summary
The property of mixed selectivity has been discussed at a computational level and offers a strategy to maximize computational power by adding versatility to the functional role of each neuron. Here, we offer a biologically grounded implementational-level mechanistic explanation for mixed selectivity in neural circuits. We define pure, linear, and nonlinear mixed selectivity and discuss how these response properties can be obtained in simple neural circuits. Neurons that respond to multiple, statistically independent variables display mixed selectivity. If their activity can be expressed as a weighted sum, then they exhibit linear mixed selectivity; otherwise, they exhibit nonlinear mixed selectivity. Neural representations based on diverse nonlinear mixed selectivity are high dimensional; hence, they confer enormous flexibility to a simple downstream readout neural circuit. However, a simple neural circuit cannot possibly encode all possible mixtures of variables simultaneously, as this would require a combinatorially large number of mixed selectivity neurons. Gating mechanisms like oscillations and neuromodulation can solve this problem by dynamically selecting which variables are mixed and transmitted to the readout.
混合选择性的性质已经在计算层面上进行了讨论,并提供了一种通过增加每个神经元功能角色的多功能性来最大化计算能力的策略。在这里,我们为神经回路中的混合选择性提供了基于生物学的implementational-level机械解释。我们定义了纯、线性和非线性的混合选择性,并讨论了如何在简单的神经回路中获得这些响应属性。对多个统计独立变量做出反应的神经元显示出混合选择性。如果它们的活动可以表示为加权和,那么它们表现出线性混合选择性;否则,它们表现出非线性混合选择性。基于不同非线性混合选择性的神经表示是高维的;因此,它们对简单的下游读出神经回路交换意见的巨大灵活性。然而,简单的神经回路不可能同时编码所有可能的变量混合,因为这需要组合大量的混合选择性神经元。振荡和神经调节等门控机制可以通过动态选择混合哪些变量并将其传输到读数来解决这个问题。
Overview
Not all brain functions are complex. They do not need to be. Simple functions can be performed by simple architectures or single layers. Seeing an object approaching, tasting a poison, detecting food in your esophagus and swallowing it, and recoiling from something that causes tissue damage are all simple functions that simple circuits and cells can accomplish. The architectures that underpin these functions can have straightforward properties or simple combinations of properties that serve this function, much like a railroad track providing a straightforward, direct route. The lack of flexibility makes these functions quick, efficient, and stereotyped.
并非所有的大脑功能都是复杂的。它们不需要复杂。简单的功能可以通过简单的架构或单层来执行。看到一个物体靠近,品尝毒药,检测食道中的食物并吞下它,从导致组织损伤的东西中退缩,这些都是简单的电路和细胞可以完成的简单功能。支撑这些功能的架构可以具有简单的属性或服务于该功能的简单属性组合,就像提供直接、直接路线的铁轨一样。缺乏灵活性使这些功能快速、高效和定型。
By contrast, the neural systems responsible for complex thought and behavior mandate flexibility. Intelligent thought is flexible thought. All creatures can react reflexively to the environment. But animals with more complex nervous systems can change how they behave by integrating more parameters into the decision-making process. They tailor ongoing behavior to the current situation and to an ever-shifting set of subgoals and goals. They also take into account an accumulating history of events that bias decision thresholds. This capacity for generalizing context-dependent behavior is crucial for our ability to project our behavior into the future, allowing us to make and execute plans.
The neural substrate for this flexibility can be seen in many places, but it is especially prevalent in cortical areas known to be critical for flexible behavior, such as the prefrontal cortex (PFC). Individually, PFC neurons have adaptive and multivariate response properties,这种灵活性的神经基础可以在许多地方看到,但它在已知对灵活行为至关重要的皮层区域尤其普遍,如前额叶皮层(PFC)。单独而言,PFC神经元具有适应性和多变量反应特性,
referred to as “mixed selectivity.” 称为“混合选择性”。
They wear many hats, showing different patterns of selectivity in different behavioral contexts. Information is often widely distributed across them. The signal-to-noise ratio of responses for each individual neuron is low, but decodable information for the population is high. PFC neurons have moderately high basal firing rates and high proportions of neurons responding to many stimuli—relying on mixing more diverse inputs that give each individual neuron many jobs encoding many variables. Our view of the PFC has evolved: rather than trains of thought on railroad tracks,他们戴着许多帽子,在不同的行为环境中表现出不同的选择性模式。信息通常广泛分布在他们之间。每个单独神经元的反应信噪比很低,但群体的可解码信息很高。PFC神经元具有适度高的基础放电率和高比例的神经元对许多刺激做出反应——依赖于混合更多样化的输入,给每个单独的神经元许多编码许多变量的工作。我们对PFC的看法发生了演变:而不是铁轨上的思维列车,
the PFC more closely resembles cars on the road and highway system. Neurons and their axons provide the highways—the anatomical architecture over which thoughts, feelings, sensations, and motor commands can travel. But the way the roads are used is vastly different based on the immense complexity of the electrical and chemical impulses being trafficked all around the brain. Vehicles of information can take different paths and different destinations. They might share a common path with some vehicles at some times and others at other times. This autonomy provides maximal freedom and dimensionality.PFC更像道路和高速公路系统中的汽车。神经元及其轴突提供了高速公路——思想、感觉、感觉和运动命令可以通过的解剖结构。但是道路的使用方式却大不相同,这是基于大脑周围传输的电和化学脉冲的巨大复杂性。信息载体可以选择不同的路径和不同的目的地。它们可能在某些时候与一些车辆共享一条共同的路径,而在其他时候与其他车辆共享一条共同的路径。这种自主性提供了最大的自由和维度。
With this comes depth of thought. The multivariate neuronal properties can increase the representational dimensionality of the population, allowing more complex computations.多变量神经元属性可以增加群体的表征维度,允许更复杂的计算。
Further, the brain tailors the dimensionality to the task at hand. Dimensionality expands and contracts to focus processing along relevant dimensions.此外,大脑根据手头的任务定制维度。维度扩展和收缩以沿着相关维度集中处理。
This keeps processing on-task and goal directed.这保持了任务和目标导向的处理。
In short, mixed selectivity gives the brain the processing power needed for complexity and flexibility. The cost of this opportunity for flexibility is the inability to take advantage of the regularities of the world to generalize to novel situations. Mixed selectivity allows representation of a large number of different situations in every detail, but sometimes we need to discard or ignore some information to make the right decision.
简而言之,混合选择性赋予大脑复杂性和灵活性所需的处理能力。这种灵活性机会的代价是无法利用世界的规律来概括到新的情况。混合选择性允许在每个细节中代表大量不同的情况,但有时我们需要丢弃或忽略一些信息来做出正确的决定。
In this perspective piece, we consider the impact of mixed selectivity on our understanding of neural processing, with the goal of grounding it in a biological implementation. To maximize the transparency of the concept of mixed selectivity, we will first define and describe mixed selectivity and then discuss its importance.
在这篇透视图中,我们考虑了混合选择性对我们理解神经处理的影响,目标是将其植根于生物学实现。为了最大限度地提高混合选择性概念的透明度,我们将首先定义和描述混合选择性,然后讨论其重要性。
What is mixed selectivity?
什么是混合选择性?
There has been some confusion around this term, which has emerged from a number of related concepts (围绕这个术语有一些混淆,它来自许多相关的概念 but the definition is simple. At its core, mixed selectivity involves a single cell showing consistent activity, which is modulated by multiple statistically independent variables. This single-cell behavior eventually has broad implications for the activity of neuronal ensembles, enabling them to process and integrate a range of independent inputs.),但定义很简单。在其核心,混合选择性涉及显示一致活动的单个细胞,由多个统计自变量调制。这种单细胞行为最终对神经元集合的活动具有广泛的影响,使它们能够处理和整合一系列独立的输入。
The history of the theory of mixed selectivity 混合选择性理论的历史
.
Since the time that artificial neural networks were conceived, their typical units have always performed two basic operations: sum multiple signals coming from different neurons and then compute a nonlinear function to this sum. It is then not surprising that the units of these networks exhibit mixed selectivity, as it is almost unavoidable if the input neurons encode different signals. What is less obvious is an understanding of the computational role of mixed selectivity neurons. This has always been one of the important roles of theoretical neuroscience. From the early days of neural network theory, it has been clear that some representations simply do not work and need to be changed. David Marr often speaks about recoding,8 invoking it when patterns of activities that need to be discriminated are too similar. The first one to realize the importance of mixing is probably Rosenblatt,9 the father of modern learning neural networks. His perceptron, in the original version (not in one discussed in the book Perceptrons10), had an intermediate layer of neurons that were randomly connected to the inputs. These neurons had mixed selectivity, and they were necessary to make the representations linearly separable and hence classifiable by a simple linear readout. Other more recent recurrent neural networks were using the same ideas (random projections) to generate mixed selectivity representations with higher dimensionality11 (echo state machine12, liquid state machine13). The idea behind the SVMs (support vector machines) with nonlinear kernels14 is again the same: to implicitly transform the representations to make them linearly separable. SVMs with nonlinear kernels are equivalent to a simple neural network with a very large intermediate layer of nonlinear units that mix the inputs nonlinearly, expanding the dimensionality. When we studied attractor neural networks that implement finite-state machines,3 we realized that we had to use a similar approach and, for the first time, we related dimensionality expansion and nonlinear mixed selectivity in clear terms.
自人工神经网络的构想以来,它们典型的单元一直执行两个基本操作:对来自不同神经元的多个信号进行求和,然后对该求和结果进行非线性函数计算。因此,如果输入神经元编码不同的信号,网络的单元呈现混合敏感性并不令人意外。较不明显的是对混合敏感性神经元的计算作用的理解。这始终是理论神经科学的重要角色之一。从神经网络理论的早期就明确了一些表示根本行不通并需要改变的事实。大卫·马尔(David Marr)经常谈到"重新编码",当需要区分的活动模式太相似时,他会提及这一概念。最早意识到混合敏感性的重要性可能是现代学习神经网络之父罗森布拉特(Rosenblatt)。他的感知器在原版(而非《感知器》一书中讨论的版本)中有一个与输入随机连接的中间层神经元。这些神经元具有混合敏感性,并且它们对于使表示线性可分、从而能够被简单线性读出所必需。其他更近期的循环神经网络也使用相同的思想(随机投影)生成具有更高维度的混合敏感性表示(如回声状态机、液态状态机)。SVMs(支持向量机)使用非线性核的思想也是相同的:隐式地转换表示,使其线性可分。带有非线性核的SVM等效于具有非常大的中间层非线性单元的简单神经网络,这些单元以非线性方式混合输入,扩展维度。当我们研究实现有限状态机的吸引子神经网络时,我们意识到我们必须采用类似的方法,并首次明确地将维度扩展和非线性混合敏感性联系在一起。
.
Besides this important but also basic general idea of dimensionality expansion, there were several other theoretical works discussing the importance of mixing in specific problems. For example, in discussions of parallel distributed processing and specifically distributed representations, the point of conjuctive coding is highly relevant to mixed selectivity. Here, Rumelhart et al. point out that the “binding problem” can be solved using neurons capable of local tuning—a capability potentially endowed by differential neuromodulation at each dendritic segment that may be electrically compartmentalized.
除了这一重要但也是基本的维数扩展一般思想之外,还有其他几部理论著作讨论了混合在特定问题中的重要性。例如,在并行分布式处理和特定分布式表示的讨论中,变相编码的点与混合选择性高度相关。在这里,Rumelhart等人指出,“绑定问题”可以使用能够进行局部调整的神经元来解决-在每个可能被电分隔的树突节段的差异神经调节可能赋予的能力。
Another example: mixed selectivity to the retinal location of a visual stimulus and the position of the eyes can be used to generate a representation of an object’s position and then determine the changes in joint coordinates needed to reach the object.另一个例子:对视觉刺激的视网膜位置和眼睛位置的混合选择性可用于生成对象位置的表示,然后确定到达对象所需的关节坐标的变化。
Neurons with mixed selectivity to the identity of a visual stimulus and its ordinal position in a sequence have been used to model serial working memory.对视觉刺激的身份及其在序列中的顺序位置具有混合选择性的神经元已被用于模拟串行工作记忆。
Mixed selectivity to stimulus identity and to a context signal have been used to model visuomotor remapping.刺激身份和上下文信号的混合选择性已被用于模拟视觉运动重映射。
More generally, complex nonlinear functions of the sensory inputs, like motor commands, can be expressed as a linear combination of basis functions.更一般地说,感觉输入的复杂非线性函数,如运动命令,可以表示为基函数的线性组合。
Pure selectivity
纯选择性
In many ways, the notion of pure selectivity can be traced to Hubel and Wiesel’s description of the “simple and complex cells” in the primary visual cortex. This pioneering work was a leap forward as it was one of the first demonstrations of the functionality of single neurons. Simple and complex cells spiked to single features. For some time, most theories of cortical function considered pure selectivity.
在许多方面,纯选择性的概念可以追溯到胡贝尔和维塞尔对初级视觉皮层中“简单和复杂细胞”的描述。这项开创性的工作是一个飞跃,因为它是单个神经元功能的首批演示之一。简单和复杂的细胞达到单一特征。一段时间以来,大多数皮层功能理论都考虑纯选择性。
The focus then was, understandably, on the first-order questions and parameters we could confidently measure—how neurons represent simple sensory features or produce motor outputs. This, after all, was one of our first forays into how cortical neurons process information. This was studied in experimental paradigms designed to be high fidelity, robust, and intentionally redundant. The purpose was to reduce variability (“noise”) so that we could be confident about the processes that sense and move our bodies within our environments.
可以理解的是,当时的重点是我们可以自信测量的一阶问题和参数——神经元如何代表简单的感官特征或产生运动输出。毕竟,这是我们对皮层神经元如何处理信息的首批尝试之一。这是在设计为高保真、稳健和有意冗余的实验范式中进行的研究。目的是减少可变性(“噪音”),这样我们就可以对在环境中感知和移动我们身体的过程充满信心。
As we built on this foundational work, our view of cortical neurons has become more dexterous and multifunctional. The more we learn, the more noise is a misnomer. It is rather factors that experimentalists did not or could not measure.随着我们在这项基础工作的基础上再接再厉,我们对皮层神经元的看法变得更加灵巧和多功能。我们学得越多,噪音就越多是用词不当。这是实验学家没有或无法测量的因素。
Initially, the idea that neurons could do more than one thing was blasphemy (one of us was accused of “turning cortex into a bowl of porridge” at a major meeting). But evidence kept amassing for multifunctionality, including observations such as neurons releasing different neurotransmitters for different functions.起初,神经元可以做不止一件事的想法是亵渎神明(我们中的一个人被指控在一次重要会议上“把皮层变成了一碗粥”)。但是关于多功能的证据不断积累,包括神经元为不同的功能释放不同的神经递质等观察。
It is now evident that pure selectivity, as important as it is, is more of an exception than the rule, especially in the cortex. Its habitats tend to be at the input edges of sensory systems and at the output edges of motor systems. In much of the rest of the cortex, the preponderance of neurons show multifunctional selectivity.现在很明显,纯粹的选择性,与它同样重要******更多的是例外而不是规则,尤其是在皮层。它的栖息地往往位于感觉系统的输入边缘和运动系统的输出边缘。在皮层的大部分其他部分,神经元的优势显示出多功能选择性。
Linear mixed selectivity
线性混合选择性
Within mixed selective neurons, there are either nonlinear or linear mixed selectivity neurons. Linear mixed selectivity在混合选择性神经元中,存在非线性或线性混合选择性神经元。
neurons are typically the result of a process of abstraction and can facilitate generalization.神经元通常是抽象过程的结果,可以促进泛化。
Linear mixed selectivity neurons show activity that can be expressed as a weighted sum of the responses to each variable (线性混合选择性神经元显示的活动可以表示为对每个变量的反应的加权和(Figure 1图1B). Consider, for instance, a neuron that responds to both written praise in an email and oral praise.B)。例如,考虑一个神经元,它对电子邮件中的书面表扬和口头表扬都有反应。
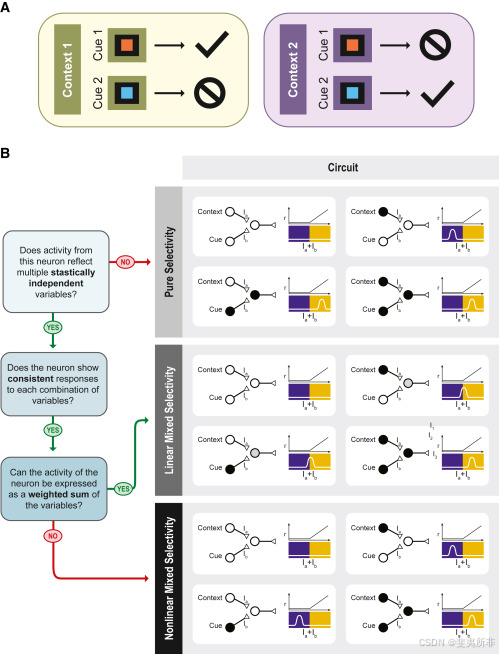
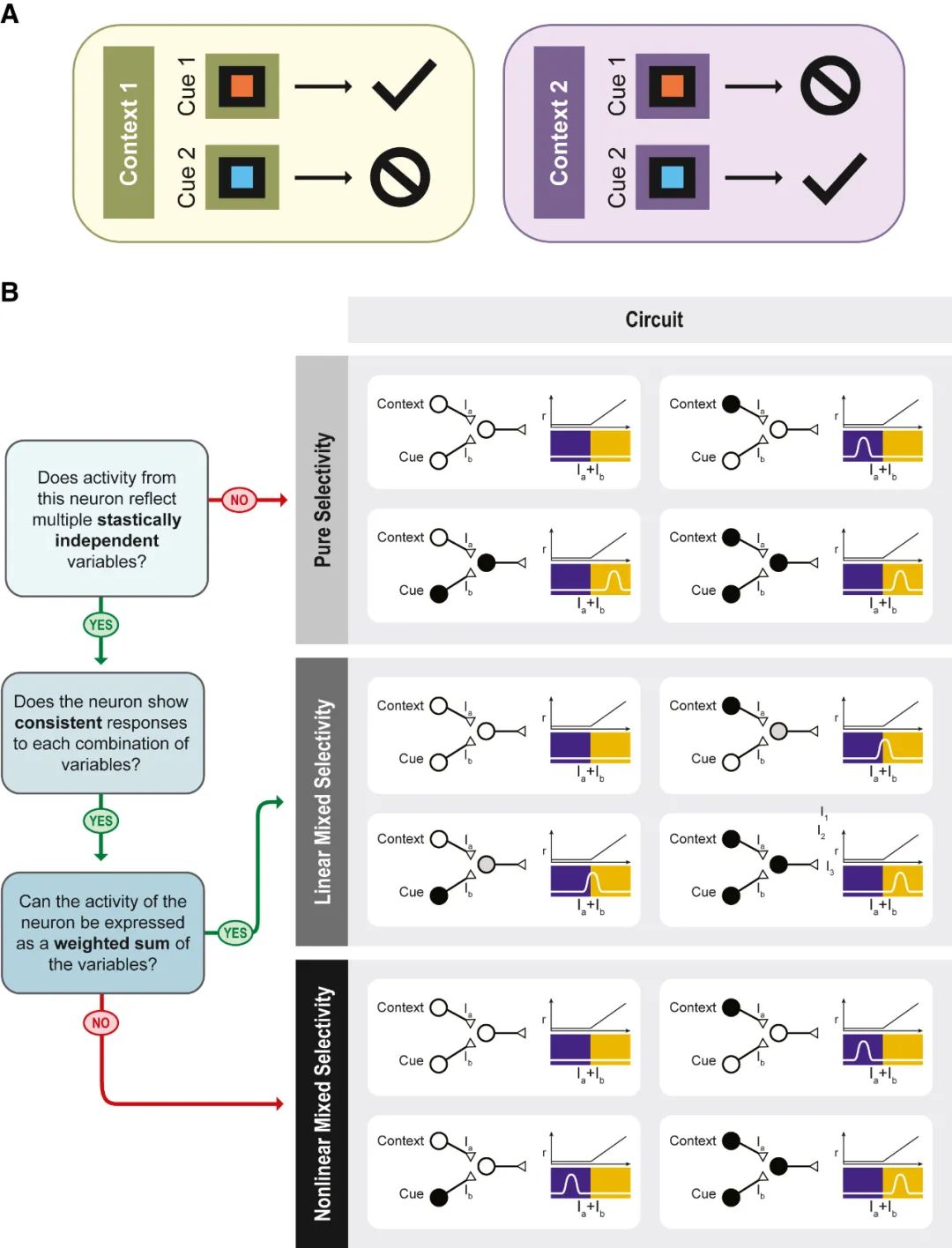
Figure 1Delineating pure, linear mixed, and nonlinear mixed selectivity using a context-dependent task
图1使用上下文相关任务描述纯、线性混合和非线性混合选择性
How does this happen? How do neurons that respond to reading and listening to praise find their way to mutually synapse on the same cell? The likely explanation is that many cells have many inputs, and those associated with environmental information that have an eventual impact on our survival, including acceptance by our social group, will be strengthened, while less significant ones will be weakened or pruned. Moreover, the surviving connections are tuned in such a way that the neuron operates in a linear regime.
这是怎么发生的?对阅读和听赞美有反应的神经元是如何在同一个细胞上找到相互突触的方式的?可能的解释是,许多细胞有许多输入,那些与最终影响我们生存的环境信息相关的信息,包括我们社会群体的接受,将得到加强,而不太重要的将被削弱或修剪。此外,幸存的连接以这样一种方式调整,即神经元以线性方式运行。
Nonlinear mixed selectivity
非线性混合选择性
Neurons that respond to multiple, statistically independent variables, where the activity tracking these two variables cannot be expressed as a weighted sum, are considered to be nonlinear mixed selective neurons (对多个统计自变量做出反应的神经元,其中跟踪这两个变量的活动不能表示为加权和,被认为是非线性混合选择性神经元(Figure 1图1B). These neurons can be modeled as computing a weighted sum of the activity of their presynaptic neurons and then passing this sum through a nonlinearity. The input neurons must encode different variables, and they can do so in multiple ways: they can be pure selectivity, nonlinear, or linear mixed selectivity neurons. Moreover, any of the numerous nonlinear mechanisms that are involved in the normal functioning of a neuron can contribute to generating nonlinear mixed selectivity: the frequency-input (f-I) curve of a neuron is typically nonlinear,B)。这些神经元可以被建模为计算其突触前神经元活动的加权和,然后通过非线性传递这个和。输入神经元必须编码不同的变量,它们可以通过多种方式做到这一点:它们可以是纯选择性、非线性或线性混合选择性神经元。此外,神经元正常功能中涉及的众多非线性机制中的任何一种都有助于产生非线性混合选择性:神经元的频率输入(f-I)曲线通常是非线性的,
and that would be sufficient. However, there are also nonlinearities in the dendritic integration and in synaptic transmission.这就足够了。然而,树枝状积分中也存在非线性并在突触传递中。
Rather than abstraction, nonlinear mixed selective neurons offer the capacity for complexity and depth. Because of their versatility, these neurons require a wide array of inputs. To have neurons be readily repurposed in different contexts also suggest a hierarchical organization. Timescales, locations, probabilities, and internal states are all variables that can contribute to context. Context-modulation of cue responding is one of the key applications of this capability. Cues are presented on discrete timescales, while contexts are represented on a longer timescale. Different contexts can influence the value or meaning of cues and, therefore, the same cue should have distinct neural representations under different contexts, as this would be the only way to produce outputs that would yield different behaviors. For example, imagine a mouse encountering an unfamiliar type of berry in its environment. If the mouse is alone and has never experienced this particular berry, it might be hesitant to eat it due to the potential risks of poisoning or illness. However, if the mouse then encounters a conspecific whose breath carries the scent of that same berry, it may interpret this as a sign that the berry is safe to consume. In this context, the smell of the berry on the breath of a fellow mouse serves as a form of social verification, indicating that the food is likely safe and worth the effort to eat. This social cue effectively shifts the mouse’s evaluation of the unfamiliar berry from potentially risky to likely safe, all based on the social transmission of food preference
非线性混合选择性神经元不是抽象的,而是提供了复杂性和深度的能力。由于它们的多功能性,这些神经元需要广泛的输入。让神经元在不同的环境中容易被重新利用也暗示了分层组织。时间尺度、位置、概率和内部状态都是有助于上下文的变量。线索响应的上下文调制是这种能力的关键应用之一。线索在离散的时间尺度上呈现,而上下文在更长的时间尺度上呈现。不同的上下文会影响线索的价值或意义,因此,同一线索在不同的上下文下应该有不同的神经表征,因为这是产生产生不同行为的输出的唯一方法。例如,想象一只老鼠在它的环境中遇到一种不熟悉的浆果。如果老鼠独自一人,从未吃过这种特殊的浆果,由于潜在的中毒或疾病风险,它可能会犹豫是否要吃它。然而,如果老鼠随后遇到一个呼吸带有同一浆果气味的同种生物,它可能会将此解释为浆果可以安全食用的标志。在这种情况下,老鼠呼吸中浆果的气味是一种社会验证形式,表明食物可能是安全的,值得努力食用。这种社会提示有效地将老鼠对不熟悉浆果的评价从潜在的风险转变为可能的安全,所有这些都基于食物偏好的社会传播
Why mix nonlinearly?
为什么要非线性混合?
Representations based on nonlinear mixed selectivity have specific computational properties: in particular, they can be high-dimensional and hence usable by a simple linear readout like a downstream neuron. These high-dimensional representations confer enormous flexibility to the neural circuit that reads them out. To understand them, it is instructive to consider what is called representational geometry (基于非线性混合选择性的表示具有特定的计算特性:特别是,它们可以是高维的,因此可以通过像下游神经元一样的简单线性读出来使用。这些高维表示与读出它们的神经回路交换意见的巨大灵活性。为了理解它们,考虑所谓的表示几何(Figure 2图2). It is easier to start from the representations that do not contain mixed selectivity: say, for example, that a visual stimulus can appear in one of two possible contexts. We denote the stimulus by S and the context by C. Both variables have only two values as there are only two stimuli and two contexts. Imagine that all the neurons have pure selectivity to either S or C. For example, we consider three neurons: the firing rate of the first one, r)。从不包含混合选择性的表示开始更容易:例如,视觉刺激可以出现在两种可能的上下文中的一种。我们用S表示刺激,用C表示上下文。两个变量都只有两个值,因为只有两个刺激和两个上下文。想象所有的神经元对S或C都有纯粹的选择性。例如,我们考虑三个神经元:第一个神经元的放电率,r11, is equal to S (pure selectivity to the stimulus); the second one, r,等于S(对刺激的纯选择性);第二个,r22, responds only to context C; and the activity of the third one, r,只响应上下文C;和第三个的活动,r33, depends on S only, but in a more complicated way, i.e., r,只依赖于S,但以更复杂的方式,即r33 = 1−S. We now consider the activity space in which r=1−S.我们现在考虑r在其中的活动空间11, r,r22, and r,和r33 are represented along the coordinate axes. In this simple example, we consider only three neurons, but typically this is a high-dimensional space with many axes. Each point in this space represents the population response in one experimental condition (e.g., the first stimulus is presented in the second context). We will have four points in the activity space as we have two contexts and two stimuli.沿着坐标轴表示。在这个简单的例子中,我们只考虑三个神经元,但通常这是一个有许多轴的高维空间。这个空间中的每个点代表一个实验条件下的群体反应(例如,第一个刺激在第二个上下文中呈现)。我们将在活动空间中有四个点,因为我们有两个上下文和两个刺激。
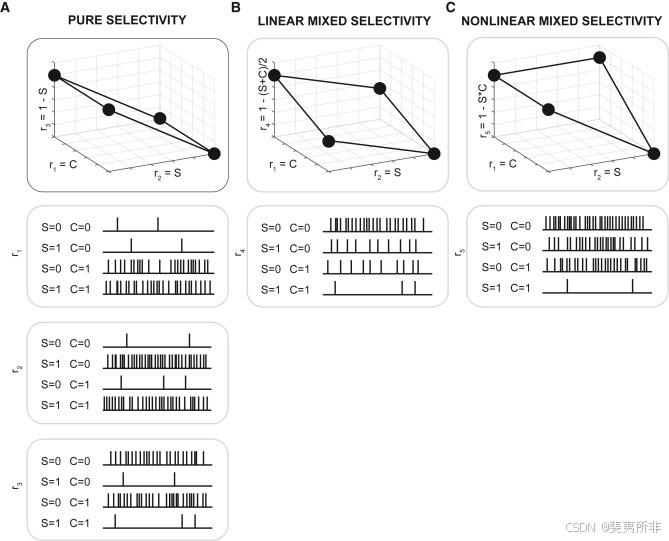
Figure 2Changes in the representational geometry due to linear and nonlinear mixed selectivity
图2 由于线性和非线性混合选择性导致的表征几何变化
The different types of selectivity yield different geometries. As the neurons have only pure selectivity, the four points will define a relatively low-dimensional object: the number of dimensions will be equal to the number of task-relevant variables. In our case, the variables are two, and the four points form a rectangle (Figure 2A). The arrangement of these points defines the geometry of the representation. In the case in which we replace the third neuron with a linear mixed selectivity neuron (Figure 2B), the geometry does not change substantially: the flat object that we saw in the case of pure selectivity neurons will rotate, but it will not change its dimensionality. The geometry changes significantly when introducing a nonlinear mixed selectivity neuron (Figure 2C). Now, the four points define a three-dimensional (3D) object called a tetrahedron.
不同类型的选择性产生不同的几何形状。由于神经元只有纯选择性,四个点将定义一个相对低维的对象:维度的数量将等于任务相关变量的数量。在我们的例子中,变量是两个,四个点形成一个矩形(图2A)。这些点的排列定义了表示的几何形状。在我们用线性混合选择性神经元替换第三个神经元的情况下(图2B),几何形状不会发生实质性变化:我们在纯选择性神经元的情况下看到的平面对象会旋转,但不会改变其维度。当引入非线性混合选择性神经元时,几何形状会发生显著变化(图2C)。现在,这四个点定义了一个称为四面体的三维物体。
Why is the dimensionality of the representation important? To understand it, we need to take the perspective of a downstream neuron reading out the representation. What this neuron can or cannot do depends on how the points of the representation are arranged. Consider, for example, the situation in which a readout neuron has to be trained to respond to a particular stimulus always in the same way, ignoring the context in which the stimulus is presented. We consider this a simple task. This can be visualized in the activity space by coloring the points according to the desired response of the readout neuron (为什么表示的维度很重要?要理解它,我们需要从一个下游神经元读出表示的角度来看。这个神经元能做什么或不能做什么取决于表示的点是如何排列的。例如,考虑一个读出神经元必须被训练以始终以相同的方式对特定刺激做出反应的情况,忽略了刺激呈现的上下文。我们认为这是一项简单的任务。这可以在活动空间中通过根据读出神经元的期望反应对点进行着色来可视化(Figure 3图3B). For example, we color purple the points corresponding to the conditions where the readout neuron should be active (S = 1) and orange the points for which the readout neuron should be inactive (S = 0).例如,我们将对应于读出神经元应该活跃的条件(S=1)的点涂成紫色,将读出神经元应该不活跃的点涂成橙色(S=0)。
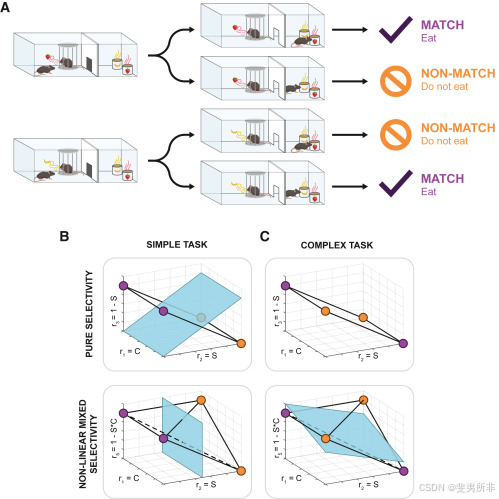
Figure 3Understanding the importance of dimensionality in neuronal readouts
图3了解维度在神经元读数中的重要性
Is it possible to connect the readout neuron to the pure selectivity neurons that we discussed in Figure 2A in such a way that it can solve this simple task? To answer this question, we need to make further simplifying assumptions. Say that the readout neuron can perform a simple operation: compute the weighted sum of the inputs and compare it with an activation threshold (linear readout). Biological neurons are complex enough to perform this operation. Graphically, we can visualize this operation by drawing a plane (Figure 3B) that separates the inputs in the activity space that activate the readout neuron (the points above the plane) from the inputs that do not reach the activation threshold (the points below the plane). For this geometry, it is possible to draw a plane separating the purple points from the orange points; in other words, a simple linear readout can perform this task. The same applies to the high-dimensional representation that involves a nonlinear mixed selectivity neuron. But consider now the same geometries when the task is more complex: the readout has to respond only to stimulus 1 when it appears in context one and to stimulus 0 when it appears in context zero, but not in the other two cases. Now, the coloring of the points is different and, interestingly, it is possible to separate the purple from the orange points in the case of the high-dimensional representation, but not in the case of the low-dimensional one (Figure 3C). This problem is equivalent to the well-known exclusive-OR (XOR) problem, and no plane can separate the points of the low-dimensional representation as required.
有没有可能将读出神经元连接到我们在图2A中讨论的纯选择性神经元,使其能够解决这个简单的任务?要回答这个问题,我们需要做进一步简化的假设。假设读出神经元可以执行一个简单的操作:计算输入的加权和,并将其与激活阈值(线性读出)进行比较。生物神经元足够复杂,可以执行这个操作。在图形上,我们可以通过绘制一个平面来可视化这个操作(图3B),它将激活读出神经元的活动空间中的输入(平面上方的点)与未达到激活阈值的输入(平面下方的点)分开。对于这个几何图形,可以画一个平面,将紫色点和橙色点分开;换句话说,一个简单的线性读数就可以完成这项任务。这同样适用于涉及非线性混合选择性神经元的高维表示。但是现在考虑同样的几何图形,当任务更复杂时:读数在上下文一中出现时只对刺激1做出反应,在上下文零中出现时只对刺激0做出反应,但在其他两种情况下没有反应。现在,点的颜色不同了,有趣的是,在高维表示的情况下可以将紫色点和橙色点分开,但在低维表示的情况下不行(图3C)。这个问题相当于众所周知的异或(XOR)问题,没有一个平面可以按照要求分离低维表示的点。
In this simple example with only four points, this coloring is the only one that does not have a linear solution. However, as the number of points increases, the colorings that require high dimensionality grow exponentially and become the majority. So, the more complex the task, the more important nonlinear mixed selectivity becomes. Note also that when the number of points increases, the maximal dimensionality increases as well. If one wants to achieve maximal dimensionality, nonlinear mixed selectivity is not sufficient, we also need an additional ingredient: the responses of the neurons have to be diverse enough. This is possible only when neurons respond to different combinations of the task-relevant variables. Maximal dimensionality confers a great flexibility to simple linear readouts, as they can separate the points in any arbitrary way or perform many different tasks. However, the maximal dimensionality can only be achieved when neurons mix nonlinearly and have diverse responses.
在这个只有四个点的简单例子中,这种着色是唯一没有线性解的。然而,随着点数的增加,需要高维度的着色呈指数增长并成为大多数。因此,任务越复杂,非线性混合选择性就越重要。还要注意,当点数增加时,最大维度也会增加。如果一个人想要达到最大维度,非线性混合选择性是不够的,我们还需要一个额外的成分:神经元的反应必须足够多样化。只有当神经元对任务相关变量的不同组合做出反应时,这才是可能的。最大维度赋予简单的线性读数极大的灵活性,因为它们可以以任何任意方式分离点或执行许多不同的任务。然而,只有当神经元非线性混合并具有不同的反应时,才能实现最大维数。
Diversity does not necessarily mean “completely unstructured.” If neuronal responses are characterized by a vector of regression coefficients, and one plots these vectors as points in a “selectivity” space, we often do not see any structure and the neurons seem to respond to random combinations of the task variables.多样性并不一定意味着“完全非结构化”如果神经元反应以回归系数向量为特征,并将这些向量绘制为“选择性”空间中的点,我们通常看不到任何结构,神经元似乎对任务变量的随机组合做出反应。
However, there are also experiments in which it is possible to observe some structure in the form of clustering: there are groups of neurons that tend to respond in a similar way.然而,也有实验可以观察到聚类形式的某种结构:有一组神经元倾向于以类似的方式做出反应。
Single-neuron versus population-coding properties
单神经元与种群编码特性
The above classification into pure, linear, or nonlinear mixed selectivity categories is done at the level of single neurons. Crucially, this first requires us to decide on a set of relevant external variables (describing, e.g., stimuli or behaviors) that we are interested in—for a different choice of this set of variables the classification of a neuron will differ in general. Let us consider again the earlier example of a neuron that responds to both written and verbal praise. If this neuron’s activity corresponds to the total amount of praise received, regardless of its mode of communication, then it could be said to exhibit pure selectivity for praise in general rather than mixed selectivity for written and verbal versions of it, even though the latter description is also valid.
以上对纯、线性或非线性混合选择性类别的分类是在单个神经元的水平上完成的。至关重要的是,这首先要求我们决定一组我们感兴趣的相关外部变量(描述,例如,刺激或行为)——对于这组变量的不同选择,神经元的分类通常会有所不同。让我们再次考虑一个神经元对书面和口头赞美都有反应的早期例子。如果这个神经元的活动对应于收到的赞美总量,不管它的交流方式如何,那么可以说它对一般赞美表现出纯粹的选择性,而不是对书面和口头版本的混合选择性,即使后一种描述也是有效的。
We can establish the selectivity properties of a single neuron by fitting a linear regression model to predict its firing rate from the external variables that we have chosen to investigate. If the fitted coefficient of only one variable is significantly different from zero, we can describe the neuron as having pure selectivity for that variable, whereas multiple non-zero coefficients would correspond to linear mixed selectivity. Furthermore, we can generalize the regression model by including features that are nonlinear combinations (e.g., pairwise products) of the external variables of interest. If this extended model can achieve a significantly higher (cross-validated) accuracy for predicting the firing rate of the neuron by assigning non-zero coefficients to these nonlinear features, the neuron can be said to exhibit nonlinear mixed selectivity. Such a regression analysis is closely related to ANOVA.我们可以通过拟合线性回归模型来建立单个神经元的选择性属性,以从我们选择研究的外部变量中预测其放电率。如果只有一个变量的拟合系数与零显著不同,我们可以将神经元描述为对该变量具有纯选择性,而多个非零系数将对应于线性混合选择性。此外,我们可以通过包括感兴趣的外部变量的非线性组合(例如成对乘积)的特征来推广回归模型。如果这个扩展模型可以通过将非零系数分配给这些非线性特征来实现预测神经元放电率的显着更高(交叉验证)的准确性,则可以说神经元表现出非线性混合选择性。这样的回归分析与方差分析密切相关。
However, beyond these single-neuron properties, we can also examine the neural representations that the activity patterns of these neurons create at the population level, and many of their properties and computational implications do not depend on the way we choose the external variables of interest. As discussed above, one particularly important geometric property of such a representation is its dimensionality. Pure and linear mixed selectivity lead to low dimensionality, equal to the number of relevant variables. Nonlinear mixed selectivity can increase the dimensionality of the representation beyond this value.
然而,除了这些单神经元属性之外,我们还可以检查这些神经元的活动模式在总体水平上创建的神经表征,它们的许多属性和计算含义并不依赖于我们选择感兴趣的外部变量的方式。如上所述,这种表征的一个特别重要的几何属性是它的维数。纯线性混合选择性导致维数较低,等于相关变量的数量。非线性混合选择性可以将表征的维数增加到该值之外。
Many computational problems can only be solved by neural networks with hidden units, i.e., they require nonlinear mixing of inputs. A network that has learned to correctly execute such a task will therefore contain at least some neurons that exhibit nonlinear mixed selectivity with respect to the variables encoded in the network inputs. However, if the network was trained for executing a particular task, it may only require very specific nonlinear mixed selectivity neurons that combine task-relevant variables in a certain way that supports the chosen task rather than a variety of different nonlinear mixed selectivity neurons with a diverse set of coding properties. In other words, even if the network performs a complex task that requires nonlinear mixing, its representations do not necessarily have to be very high dimensional.
许多计算问题只能通过具有隐藏单元的神经网络来解决,即它们需要输入的非线性混合。已经学会正确执行这样的任务的网络因此将包含至少一些相对于网络输入中编码的变量表现出非线性混合选择性的神经元。但是,如果网络被训练用于执行特定任务,它可能只需要非常特定的非线性混合选择性神经元,这些神经元以某种支持所选任务的方式组合任务相关变量,而不是各种不同的非线性混合选择性神经元,具有不同的编码属性集。换句话说,即使网络执行需要非线性混合的复杂任务,其表示也不一定必须是非常高维的。
However, representations with (close to) maximal dimensionality, which are achieved only when the responses of different neurons are sufficiently diverse, are an important coding scheme in situations in which a population of neurons does not have sufficient information about which task it is meant to support (perhaps because the neurons are not provided with rich and individually tailored error signals as they would be, e.g., during learning via backpropagation) or if the animal needs to potentially execute a large class of different tasks and flexibly switch between them. In such scenarios, the neural population cannot shape its representations to specifically subserve a particular task, but it can nevertheless try to form a neural representation of the task-relevant variables that is generally useful for executing many possible tasks. One concrete way to achieve this is to create a high-dimensional representation that enables a downstream neuron implementing a linear readout (or linear classifier) to correctly extract many functions of the input (perhaps even all possible dichotomies for a moderate number of binary inputs). In this sense, high dimensionality is associated with the (cognitive) flexibility of the animal—performing new tasks is then simply a matter of finding a different readout, which can be done using a simple perceptron or delta rule, but does not require changing the internal representations, which would necessitate more sophisticated learning algorithms.
然而,只有当不同神经元的反应足够多样化时,具有(接近)最大维度的表示才会实现,在神经元群体没有足够的信息来说明它应该支持哪项任务的情况下,这是一种重要的编码方案(也许是因为神经元没有像它们那样提供丰富的和单独定制的错误信号,例如,在通过反向传播学习期间),或者如果动物需要潜在地执行一大类不同的任务并在它们之间灵活切换。在这种情况下,神经群体无法塑造其表示以专门支持特定任务,但它仍然可以尝试形成任务相关变量的神经表示,这通常对执行许多可能的任务有用。实现这一目标的一个具体方法是创建一个高维表示,使实现线性读出(或线性分类器)的下游神经元能够正确提取输入的许多功能(甚至可能是中等数量二进制输入的所有可能二分法)。从这个意义上说,高维与动物的(认知)灵活性有关——执行新任务只是找到不同的读出,这可以使用简单的感知器或增量规则来完成,但不需要改变内部表示,这将需要更复杂的学习算法。
Expression of mixed selectivity across circuits and structures
跨电路和结构的混合选择性的表达式
To appreciate what our brains might look like without mixed selectivity, we can examine which functions and structures at the micro- and macrocircuit level rely on mixed selectivity. Few functions are completely devoid of circuit flexibility, and although the labeled lines circuit motif is the oldest in neuroscience, it is not the most prevalent.为了理解我们的大脑在没有混合选择性的情况下会是什么样子,我们可以检查微观和宏观电路水平上的哪些功能和结构依赖于混合选择性。很少有功能完全没有电路灵活性,尽管标记线电路母题在神经科学中是最古老的,但它并不是最普遍的。
Conversely, mixed selectivity was discovered in the context of higher cognitive functions, allowing the brain to learn new rules and switch between different rules in different contexts.相反,在高级认知功能的背景下发现了混合选择性,允许大脑学习新规则并在不同背景下的不同规则之间切换。
These computations for complexity can be seen in cells and circuits down to the olfactory glomeruli and in the auditory and somatosensory cortex. 这些计算的复杂性可以在细胞和回路中看到,一直到嗅肾小球。在听觉中和体感皮层。
The fact that we can learn to like poisons (like caffeine and alcohol) and learn to hate calorie-rich foods (pistachio ice cream after food poisoning) suggests that even our sensory systems have flexibility.事实上,我们可以学会喜欢毒药(如咖啡因和酒精)和讨厌富含卡路里的食物(食物中毒后的开心果冰淇淋),这表明即使我们的感官系统也有灵活性。
There are many circuit motifs that can give rise to flexibility, circuits that diverge depending on gating, that converge to be integrated, or that compete to orchestrate competing mechanisms,有许多电路图案可以产生灵活性,电路根据门控而分化,汇聚成集成,或者竞争协调竞争机制,
and the principles of mixed selectivity are foundational to them all.混合选择性原则是它们的基础。
Labeled lines circuits have their perks. Signal-to-noise ratio is maximized, as crosstalk is minimized. The lack of crosstalk guarantees high fidelity. Moreover, pure selectivity neurons can have other advantages in terms of energy consumption and number of needed connections. Although the representations based on pure and linear mixed selectivity are completely equivalent from the point of view of a linear readout (they have the same geometry), when one imposes that the neural activity can only be positive, the pure selectivity neurons consume less energy.标记线电路有其特殊之处。信噪比最大化,因为串扰最小化。串扰的缺乏保证了高保真。此外,纯选择性神经元在能量消耗和所需连接数量方面可以有其他优势。尽管从线性读数的角度来看,基于纯和线性混合选择性的表示完全等价(它们具有相同的几何形状),但当一个人强加神经活动只能是正的时,纯选择性神经元消耗的能量更少。
Let us begin by considering sensory association systems wherein the goal is to produce appropriate reliable responses to stimuli. Of all types of information, getting reliable sensory information about our dynamic environment as we navigate it is paramount. Bipolar cells relaying information from photoreceptors to retinal ganglion cells do not require complexity or depth as much as fidelity—they know what information that photoreceptor is providing, which makes decoding trivial.
让我们首先考虑感官关联系统,其目标是对刺激产生适当可靠的反应。在所有类型的信息中,在我们导航时获得关于我们动态环境的可靠感官信息至关重要。将信息从光感受器传递到视网膜神经节细胞的双极细胞不需要复杂性或深度,而是需要保真度——它们知道光感受器提供了什么信息,这使得解码变得微不足道。
Initially, our brains filter incoming sensory information, forwarding for full processing only what is important to send along. Neurons within the thalamic nuclei handle this initial filtration, sending massive spikes that increase ∼500%最初,我们的大脑过滤传入的感官信息,只转发重要的信息进行全面处理。丘脑核内的神经元处理这种初始过滤,发送大量增加约500%的尖峰
from their basal activity for the sharpest, most fleeting of signals to be unambiguously broadcast to multiple distributed systems. In parallel, other sensory processing systems through the auditory cortex, for example, will undergo plasticity based on experience, behavioral state, hormonal signals, etc.从它们最清晰的基础活动,最短暂的信号被明确地广播到多个分布式系统。与此同时,例如,通过听觉皮层的其他感官处理系统,将根据经验、行为状态、荷尔蒙信号等进行可塑性。
The thalamus sends information to many places throughout the corticolimbic system丘脑通过皮质边缘系统向许多地方发送信息
—even the basal ganglia,-甚至基底神经节,
including the basolateral amygdala (BLA) and the PFC.包括基底外侧杏仁核(BLA)和PFC。
The BLA is a hub for integrating sensory information and rapidly converting that into a behavioral response.BLA是整合感官信息并将其快速转化为行为反应的枢纽。
The amygdala is a relatively primitive structure with the capacity for plasticity to form new associations and lies in the middle of the spectrum. The amygdala has been demonstrated to have a substantial amount of hard-wiring杏仁核是一种相对原始的结构,具有形成新联想的可塑性,位于光谱的中间。杏仁核已被证明有大量的硬线
in terms of certain neurons driving specific functions irrespective of state or context, although there is still some evidence for mixed selectivity for some variables (see, e.g., O’Neill et al.就某些神经元驱动特定功能而言,无论状态或上下文如何,尽管仍然有一些证据表明某些变量具有混合选择性(参见,例如,O’Neill et al.). Further support for the notion of fixed functions existing in the BLA include work demonstrating the existence of a long-lasting engram or memory trace that is stored long term in the same cells in which it was formed.)。对BLA中存在的固定功能概念的进一步支持包括证明存在长期存在的印记或记忆痕迹的工作,这些印记或记忆痕迹长期储存在形成它的相同细胞中。
Both the BLA and PFC receive sensory information from many inputs, including the thalamus, but process it differently.BLA和PFC都从包括丘脑在内的许多输入接收感官信息,但处理方式不同。
“Structured” as a divergence point for positive and negative valence representation, the amygdala has some neurons selectively responding to positive valence and others to negative,“结构化”作为正价和负价表示的分歧点,杏仁核有一些神经元选择性地对正价做出反应,另一些神经元选择性地对负价做出反应,
though it mixes some other variables.尽管它混合了一些其他变量。
The striatum then collates these different signals, weighs them, and decides on a single motor plan from several rehearsed or innate motor sequences.纹状体然后整理这些不同的信号,权衡它们,并从几个排练或先天的运动序列中决定一个单一的运动计划。
Conversely, the PFC approaches the received information with precisely the opposite strategy as it has diametrically distinct functional goals from the sensorimotor chain. Its computational aim is to weigh choices and delve deeper rather than produce fixed motor responses. Thus, a different set of tools are needed. This requires a different cellular and synaptic architecture that deviates from hardwired functions, opting for maximal flexibility. This flexibility is achieved by sending diffuse signals everywhere, each dimension represented by a combination of decentralized signals, allowing any number of readily decipherable messages to be selected for readout at any given time.
相反,PFC以完全相反的策略处理接收到的信息,因为它与感觉运动链有着截然不同的功能目标。它的计算目标是权衡选择并深入研究,而不是产生固定的运动反应。因此,需要一套不同的工具。这需要不同于硬连线功能的细胞和突触架构,选择最大的灵活性。这种灵活性是通过向各处发送扩散信号来实现的,每个维度都由分散信号的组合表示,允许在任何给定时间选择任意数量的易于破译的信息进行读出。
Unlike the strong signals sent out by the amygdala, the PFC communicates with a hum of whispers. Classic plots of peri-stimulus time rasters or histograms show high baseline noise with modest signals —but an extraordinarily high number of variables can be decoded from PFC activity. Importantly, high dimensionality is not random in the sense that these neurons are always processing different signals; they are reliable and perform consistently within the same context, showing different responses in varying contexts. Information is not represented by individual cells but is more prevalent in the population.与杏仁核发出的强烈信号不同,PFC以低语的嗡嗡声进行交流。经典的刺激周围时间栅格图或直方图显示了高基线噪声和适度的信号——但可以从PFC活动中解码出非常多的变量。重要的是,高维度不是随机的,因为这些神经元总是在处理不同的信号;它们是可靠的,在相同的环境中表现一致,在不同的环境中表现出不同的反应。信息不是由单个细胞表示的,而是在人群中更普遍。
It is routed in different “subspaces” through population dynamics rather than individual neurons.它是通过群体动力学而不是单个神经元在不同的“子空间”中路由的。
The hippocampus and dentate gyrus utilize a sparse coding strategy, with most neurons silent at any given moment海马体和齿状回采用稀疏编码策略,大多数神经元在任何给定时刻都保持沉默
and this coding scheme densely packed into layer CA1 still involves neurons with mixed selectivity to position, head direction, and speed of an animal that freely explores an environment.这种密集包装在CA1层的编码方案仍然涉及对自由探索环境的动物的位置、头部方向和速度具有混合选择性的神经元。
Remapping can be interpreted as another expression of this coding scheme: hippocampal neurons encode sequences, the delivery of reward, and the encounter with other animals.重映射可以解释为这种编码方案的另一种表达:海马神经元编码序列、奖励的传递以及与其他动物的相遇。
All these variables affect the neural representations and they can easily result in some form of remapping when mixed together with position. This is not surprising if one assumes that the hippocampus is fundamentally a memory system that compresses the complex sensory representations by taking advantage of their regularities (see, e.g., Gluck and Myers所有这些变量都会影响神经表征,当它们与位置混合在一起时,很容易导致某种形式的重新映射。如果假设海马体从根本上说是一个记忆系统,它利用复杂的感觉表征的规律性来压缩复杂的感觉表征,这并不奇怪(参见例如Gluck和Myers and Benna and Fusi还有Benna和Fusi). The resulting representations are sparser, more decorrelated, higher dimensional, and involve mixed selectivity neurons that encode all the different aspects of sensory experiences that are memorized.)。由此产生的表征更稀疏、更去相关、更高维度,并涉及混合选择性神经元,这些神经元编码记忆的感官体验的所有不同方面。
Unlike sparse coding systems like the hippocampus, the PFC employs a more proliferative coding strategy. For instance, in the amygdala, approximately 10%–15% of neurons will respond to a salient stimulus, like a footshock-predicting stimulus, at low basal firing rates (∼1 Hz).与海马体等稀疏编码系统不同,PFC采用更具增殖性的编码策略。例如,在杏仁核中,大约10%-15%的神经元会在低基础放电率(1赫兹)下对突出刺激(如footshock-predicting刺激)做出反应。
However, in the PFC, the same predictive cue will be encoded by around 30%–40% of neurons, albeit with a lower signal-to-noise ratio (basal firing rates ∼10 Hz).然而,在PFC中,相同的预测线索将由大约30%-40%的神经元编码,尽管信噪比较低(基本放电率约10 Hz)。
These differences in coding strategies in different micro- and macrocircuits across the brain endow the brain with its versatility and may help illustrate the functional utility of mixed selectivity when it is expressed to greater or lesser degrees within the mammalian brain.
不同的微电路和宏电路在整个大脑中编码策略的这些差异赋予了大脑的多功能性,并可能有助于说明混合选择性的功能效用,当它在哺乳动物大脑中表达或多或少的程度。
Not everything can be mixed
不是所有的东西都可以混合
We often observe that neural representations supporting a specific task exhibit the maximal dimensionality enabled by the task 我们经常观察到支持特定任务的神经表示表现出任务启用的最大维度
and, hence, that the task-relevant variables are mixed in all possible ways. However, the experiments performed in a laboratory in a highly controlled environment typically involve a relatively small number of variables, sometimes only one or two. What happens when the subjects perform a real-world task? In a simple task, like the one considered in 因此,与任务相关的变量以各种可能的方式混合在一起。然而,在高度受控的环境中进行的实验室实验通常涉及相对较少数量的变量,有时只有一两个。当受试者执行现实世界的任务时会发生什么?在一个简单的任务中,就像在Figure 1图1 with only two binary variables, context, and stimulus, the total number of possible conditions is 4. In general, however, as a function of the number of binary variables V, the number of task conditions grows exponentially as 2只有两个二元变量,上下文和刺激,可能条件的总数为4。然而,一般来说,作为二元变量V数量的函数,任务条件的数量呈指数增长为2VV. For a complex, real-work task expressing a sizable number of task-relevant variables V, this number can therefore be huge, meaning that reaching maximal dimensionality would require a correspondingly large number of mixed selectivity neurons. Because then the maximal dimensionality is bounded by the number of neurons and the number of conditions,.对于表达大量任务相关变量V的复杂、实际工作任务,这个数字因此可能很大,这意味着达到最大维度将需要相应大量的混合选择性神经元。因为最大维度受神经元数量和条件数量的限制,
we might reach the upper limit imposed by the size of the neural population before all variable combinations are possibly encoded in mixed selectivity neurons.在所有变量组合可能被编码在混合选择性神经元中之前,我们可能会达到由神经群体大小施加的上限。
Fortunately, the world is highly structured and solving a realistic task typically departs from the worst-case scenario that would require mixing all variables. For example, behavioral contexts are often compartmentalized such that we might have to mix a subset of variables in one task and distinct sets of variables in different tasks but never have to worry about mixing variables that are relevant in different tasks. If we can selectively gate the variables we are interested in, then the number of required neurons would be significantly smaller. In a simple example, where 2 tasks need to be implemented and each only involves a distinct subset of V/2 variables, we would need 2幸运的是,世界是高度结构化的,解决现实任务通常会偏离需要混合所有变量的最坏情况。例如,行为上下文通常被划分为不同的部分,因此我们可能不得不在一项任务中混合变量的子集,在不同的任务中混合不同的变量集,但永远不必担心混合与不同任务相关的变量。如果我们可以选择性地选择我们感兴趣的变量,那么所需的神经元数量将大大减少。在一个简单的例子中,需要执行2个任务,每个任务只涉及V/2变量的不同子集,我们需要2个V/2V/2 + 2 + 2V/2V/2 = 2 = 2V/2+1V/2+1 neurons as opposed to 2神经元而不是2VV. The plot in .情节在Figure 4图4A shows how many neurons we need when we have to mix only a fraction, F, of the variables. As the world becomes more structured (i.e., 1/F increases), the number of needed neurons decreases very rapidly (note that the scale of the y axis is logarithmic). How can we implement such a gating mechanism that would allow our population to exploit this structure? One simple answer is to choose properly the connections and the response properties of the input neurons. Indeed, properly connected nonlinear neurons can readily implement the most general form of gating and solve arbitrarily complex problems. However, it is sometimes difficult to learn these connectivity schemes, and there are other forms of gating that can complement those based on the careful choice of the circuit architecture. For example, there are at least two other mechanisms that implement some form of dynamic gating (see A显示了当我们只需要混合一小部分变量F时,我们需要多少神经元。随着世界变得更加结构化(即1/F增加),所需神经元的数量非常迅速地减少(注意y轴的比例是对数的)。我们如何实现这样一种门控机制,让我们的群体利用这种结构?一个简单的答案是正确选择输入神经元的连接和响应属性。事实上,正确连接的非线性神经元可以很容易地实现最一般形式的门控,并解决任意复杂的问题。然而,有时很难学习这些连接方案,还有其他形式的门控可以补充那些基于仔细选择电路架构的门控。例如,至少还有另外两种机制可以实现某种形式的动态门控(参见Figure 4图4B), selecting which neurons to listen to depending on the task or the context. There are two implementational motifs for gating responses or signaling to a given neuron that it should participate in one ensemble or another: electrical and chemical. Oscillatory dynamics naturally organize neural activity into functional patterns (B),根据任务或上下文选择要听的神经元。有两种实现模式,用于门控响应或向给定神经元发出信号,表明它应该参与一个或另一个合奏:电的和化学的。振荡动力学自然地将神经活动组织成功能模式(Figure 4图4C). Neuromodulatory signals rely on the diverse distribution of receptor expression profiles to signal the appropriate ensemble to amplify depending on cues, contexts, or internal states (C)。神经调节信号依赖于受体表达谱的不同分布,根据线索、上下文或内部状态(Figure 4图4D). These two mechanisms likely work together to dynamically form functional networks.D)。这两种机制可能协同工作以动态形成功能网络。
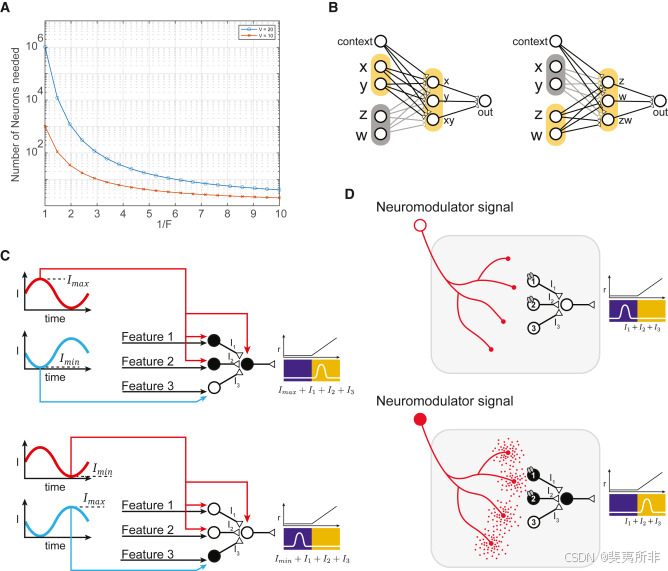
Figure 4Strategies to orchestrate gating
图4编排门控的策略
Oscillatory dynamics flexibly organize mixed selectivity neurons
振荡动力学灵活组织混合选择性神经元
Like a stadium crowd doing “the wave,” mixed selectivity neurons can be organized on-the-fly, shifting their participation in different ensembles and networks for different functions. This organization has to occur in real-time at a scale large enough to produce function. One can see such organization in the oscillations of local field potentials (LFPs).就像体育场人群做“波”一样,混合选择性神经元可以动态组织,改变它们参与不同的集合和网络以实现不同的功能。这种组织必须以足够大的规模实时发生以产生功能。人们可以在局部场电位(LFP)的振荡中看到这种组织。
They reflect coordinated changes in neuronal excitability at the mesoscale, involving millions of neurons.它们反映了中尺度神经元兴奋性的协调变化,涉及数百万个神经元。
It is at this scale that the brain focuses attention, makes decisions, executes actions, and retrieves memories—processes that necessitate the coordination of numerous neurons.正是在这种规模下,大脑集中注意力、做出决定、执行行动和检索记忆——这一过程需要众多神经元的协调。
There is ample evidence for a role for neural oscillations in the dynamic organization of functional networks. Top-down information is reflected in patterns of LFP coherence.有充分的证据表明神经振荡在功能网络的动态组织中发挥作用。自上而下的信息反映在LFP相干的模式中。
Changes in oscillatory dynamics track changes in attentional focus and state.振动动力学的变化跟踪注意焦点和状态的变化。
Oscillations help route information and form network assemblies in the hippocampus and cortex.振荡有助于传递信息并在海马和皮层形成网络组件。
Further, LFPs serve as reliable sources of information, unaffected by neuronal representation drift.此外,LFP作为可靠的信息来源,不受神经元表示漂移的影响。
Further, oscillations form traveling waves此外,振荡形成行波
that can have precise influences on networks and impact function.这可以对网络和影响函数产生精确的影响。
The theory of spatial computing ties this together to explain how the brain applies rhythms to physical patches of the cortex to selectively control just the right neurons at the right times to do the right things.空间计算理论将这些联系在一起来解释大脑如何将节奏应用于皮层的物理补丁,以在正确的时间选择性地控制正确的神经元来做正确的事情。
Importantly, the fluctuating electric fields not only reflect organization but can also create organization by having a causal influence. There are numerous instances of ephaptic coupling (i.e., the causal influence of electric fields) in the brain.重要的是,波动的电场不仅反映组织,而且还可以通过因果影响来创建组织。大脑中有许多触觉耦合(即电场的因果影响)的实例。
When cortical neurons are not spiking (which is much of the time) their membrane potentials are oscillating below the spiking threshold. This reflects and contributes to the surrounding electric fields. Thus, cortical neurons spend much of their time “teetering” on the edge of spiking in a sea of fluctuating electric fields. Even small changes in the fields can “push” them one way or another. Electric fields are an ideal “orchestra conductor” for coordinating neural activity. They spread influences at the speed of light. It would be remarkable if evolution did not take advantage of this phenomenon to use it for organizing neural activity.当皮层神经元没有尖峰时(这是大部分时间),它们的膜电位在尖峰阈值以下振荡。这反映并有助于周围的电场。因此,皮层神经元大部分时间都在波动电场的海洋中“摇摇欲坠”。即使是电场的微小变化也能以这样或那样的方式“推动”它们。电场是协调神经活动的理想“管弦乐队指挥”。它们以光速传播影响。如果进化没有利用这种现象来组织神经活动,那将是非常了不起的。
Oscillations likely interact with neuromodulation. Neurons corelease fast neurotransmitters as well as neuromodulators, which are packaged into either vesicles or dense core vesicles.振荡可能与神经调节相互作用。神经元共同释放快速神经递质和神经调质,它们被包装成囊泡或致密的核心囊泡。
Neuromodulation affects excitability. Plus, certain oscillatory frequencies can cause preferential release of dense core vesicles containing neuromodulatory signals. This offers another layer of control.神经调节影响兴奋性。此外,某些振荡频率会导致含有神经调节信号的致密核心囊泡优先释放。这提供了另一层控制。
Neuromodulatory signals tune ensemble volume, orchestrating mixed selectivity
神经调节信号调节合奏音量,编排混合选择性
Another organizational push may come from neuromodulation, which can have effects in hundreds of milliseconds and last for hours.另一个组织推动可能来自神经调节,它可以在数百毫秒内产生效果,并持续数小时。
Neuromodulation can trigger the transition between behavioral states,神经调节可以触发行为状态之间的转换,
often by activating a neuromodulatory or neuropeptidergic nucleus that can modulate the release of a neuromodulator. These neuromodulators have broad but unique innervation patterns across the brain. Their signals will be read out through axonal innervation patterns and downstream receptor expression profiles. The receptor expression profiles are predominantly predetermined but subject to experience-dependent plasticity. They can act as a weighted filter, allowing different neuromodulators to generate varied brain states. From ghrelin inducing hunger to oxytocin-stimulating prosocial behavior, neuromodulatory systems offer a wide spectrum of programs.通常通过激活神经调质或神经肽能核来调节神经调质的释放。这些神经调质在整个大脑中具有广泛但独特的神经支配模式。它们的信号将通过轴突神经支配模式和下游受体表达谱来读取。受体表达谱主要是预先确定的,但受制于experience-dependent可塑性。它们可以充当加权过滤器,允许不同的神经调质产生不同的大脑状态。从饥饿素诱导饥饿到oxytocin-stimulating亲社会行为,神经调节系统提供了广泛的程序。
There are different uses for neuromodulation and they can have specific effects. In the BLA, at baseline conditions, there is a bias toward prioritizing negative valence—which is adaptive given that predation is a more immediate threat to survival than not obtaining food or water. The relative bias can be altered or even flipped by changes in internal or external conditions. Food restriction can shift the balance between positive and negative valence processing circuits in the BLA,神经调节有不同的用途,它们可以产生特定的效果。在BLA中,在基线条件下,有优先考虑负价的偏见——这是自适应的,因为捕食比得不到食物或水对生存构成更直接的威胁。相对偏差可以通过内部或外部条件的变化来改变甚至翻转。食物限制可以改变BLA中正负价处理回路之间的平衡,
which may facilitate the prioritization of food-seeking via changes in hormonal, peptidergic, or modulatory signals. Additionally, increasing the concentration of neurotensin in the BLA gates reward learning, effectively shifting the bias toward reward learning.这可能通过荷尔蒙、肽能或调节信号的变化促进寻找食物的优先顺序。此外,增加BLA门中神经降压素的浓度奖励学习,有效地改变奖励学习的偏见。
Multivariate and specific effects of neuromodulation are evident in the PFC, where mixed selectivity is expressed by most neurons. Dopamine can serve as the “switch operator” for directing information flow.神经调节的多变量和特异性效应在PFC中很明显,其中大多数神经元表达混合选择性。多巴胺可以作为引导信息流的“开关操作员”。
In the mPFC, increasing dopaminergic tone amplifies the signal-to-noise ratio for information about aversive, but not appetitive, stimuli in a specific projection to a brainstem region, the periaqueductal gray (PAG).在mPFC中,增加的多巴胺能音调放大了脑干区域导水管周围灰度(PAG)特定投射中厌恶但非食欲刺激信息的信噪比。
Importantly, dopamine concentration does not act uniformly on different PFC neurons; mPFC neurons projecting to the nucleus accumbens (Nac) showed a suppression of activity, while mPFC-PAG neurons showed a selective amplification of information about punishments, such as air puff or foot shock, but not rewards such as sucrose.重要的是,多巴胺浓度不统一地作用于不同的PFC神经元;mPFC神经元投射到伏隔核(Nac)显示抑制活动,而mPFC-PAG神经元显示选择性放大有关惩罚的信息,如空气泡芙或脚电击,但不是奖励如蔗糖。
Importantly, dendritic nonlinearities are a key subcellular component that provide a possible mechanistic explanation for how mixed selectivity can be implemented in the context of neuromodulation. Various neuromodulatory receptors may be expressed and multiplexed on a single cell, allowing a given cell to be recruited (or suppressed and effectively excluded) from a given ensemble. Neuronal dendrites offer a high level of electrical compartmentalization onto which different functional classes of receptors (ranging from excitatory or inhibitory to different neuromodulatory receptors) may be segregated into different compartments,重要的是,树突状非线性是一个关键的亚细胞成分,它为如何在神经调节的背景下实现混合选择性提供了可能的机械解释。各种神经调节受体可以在单个细胞上表达和复用,允许从给定的集合中招募(或抑制并有效地排除)给定的细胞。神经元树突提供了高水平的电compartmentalization,不同功能类别的受体(从兴奋性或抑制性到不同的神经调节受体)可以被分离到不同的隔间,
allowing the cell to receive and transmit information differently in the presence of different concentrations of neuromodulators—such as dopamine, serotonin, and norepinephrine—as demonstrated in acetylcholine.允许细胞在不同浓度的神经调节剂(如多巴胺、血清素和去甲肾上腺素)存在下以不同的方式接收和传递信息,如乙酰胆碱所示。
For example, a single cell may have a proximal dendritic compartment receiving bottom-up sensory input and a distal dendritic compartment receiving top-down, predictive coding inputs, and allow a cell to integrate both of these signals.例如,单个细胞可以具有接收自下而上的感觉输入的近端树枝状隔室和接收自上而下的预测编码输入的远端树枝状隔室,并且允许细胞整合这两种信号。
Finally, the chemical signal of neuromodulation binds to GPCRs, initiating second-messenger cascades that can contribute to shifting cells into different membrane states, thus altering the electrical signals. By biasing resting membrane potentials, and using different receptors to induce different changes to the cell, neuromodulators can shift the synchrony and the oscillatory patterns of the brain.
最后,神经调节的化学信号与GPCRs结合,启动第二信使级联,这有助于将细胞转移到不同的膜状态,从而改变电信号。通过偏置静息膜电位,并使用不同的受体诱导细胞发生不同的变化,神经调质可以改变大脑的同步性和振荡模式。
Concluding remarks
结束语
When the first description of mixed selectivity was articulated over a decade ago, the initial focus was on the population dynamics of the PFC and a high-level cognitive function, working memory. Now, we know that mixed selectivity is not a rare feature of neurons of certain brain areas, organisms, and functions. Rather, mixed selectivity is ubiquitous.十多年前,当人们首次描述混合选择性时,最初的焦点是PFC的群体动态和高级认知功能——工作记忆。现在,我们知道混合选择性并不是某些大脑区域、生物体和功能神经元的罕见特征。相反,混合选择性无处不在。
It is present across species and across functions from high-level cognition to “automatic” sensorimotor processes such as object recognition and even to homeostatic processes.它存在于跨物种和跨功能中,从高级认知到“自动”感觉运动过程,如物体识别甚至是自我平衡过程。
The widespread presence of mixed selectivity underscores its fundamental role in providing the brain with the scalable processing power needed for complex thought and action.混合选择性的广泛存在强调了它在为大脑提供复杂思想和行动所需的可扩展处理能力方面的基本作用。
Acknowledgments
致谢
Special thanks to Amy Cao for her invaluable contributions to the design and artistic rendering of the figures. K.M.T. is an HHMI Investigator, member of the Kavli Institute for Brain and Mind, and the Wylie Vale chair at the Salk Institute for Biological Studies, and this work was supported by funding from Salk, HHMI, Kavli Foundation, Dolby Family Fund, R01-MH115920 (NIMH), R37-MH102441 (NIMH), and Pioneer Award DP1-AT009925 (NCCIH). E.K.M. was supported by the Office of Naval Research N00014-22-1-2453, NIMH 1R01MH131715-01, The JPB Foundation, and The Picower Institute for Learning and Memory. S.F. was supported by the Simons Foundation, the Gatsby Charitable Foundation, and the Swartz Foundation. M.K.B was supported by R01NS125298 (NINDS) and the Kavli Institute for Brain and Mind. M.R. was supported by the European Union’s Horizon Europe research and innovation programme under grant agreement no. 101070408 (SustainML).
特别感谢艾米·曹对人物设计和艺术渲染的宝贵贡献。K. M.T.是HHMI研究员,Kavli脑与心智研究所成员,索尔克生物研究所Wylie Vale主席,这项工作得到了索尔克、HHMI、Kavli基金会、杜比家庭基金、R01-MH115920(NIMH)、R37-MH102441(NIMH)和先锋奖DP1-AT009925(NCCIH)的资助。E.K.M.得到了海军研究办公室N00014-22-1-2453、NIMH 1R01MH131715-01、JPB基金会和皮考尔学习和记忆研究所的支持。S.F.得到了西蒙斯基金会、盖茨比慈善基金会和斯沃茨基金会的支持。M.K.B得到了R01NS125298(NINDS)和Kavli脑与心智研究所的支持。M.R.得到了欧盟Hors的支持101070408(持续ML)。
Declaration of interests
K.M.T. and E.K.M. are members of the Neuron advisory board.
via:
-
Mixed selectivity: Cellular computations for complexity: Neuron Published:May 09, 2024